
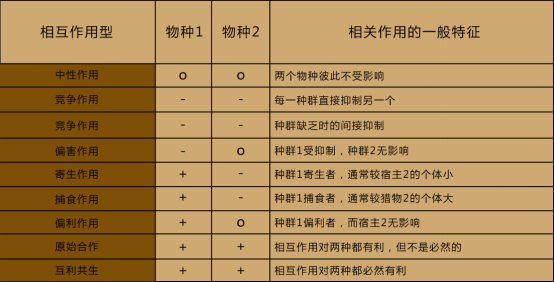
种间关系有哪些相互作用的类型呢?我们根据两个物种间的关系,把种间相互作用分为以下类型:
(1)中性作用(2)竞争作用(3)偏害作用(4)寄生作用
(5)捕食作用(6)偏利作用(7)元适合做(8)互利共生
![]()
1.种间竞争的概念:
种间竞争是指具有相似要求的不同物种,为了争夺空间和资源,而产生的一种直接或间接抑制对方的现象。
2.竞争结果:
(1)淘汰:即一个种群被另一个种群完全排挤掉;
(2)习性变化:即一个种群迫使另一种群占有不同的空间;使其食性特化或其他生态习性分化,如时间分隔等。
这也就说明,竞争能力可以决定生态习性、生活型和生态幅度等因素。
![]()
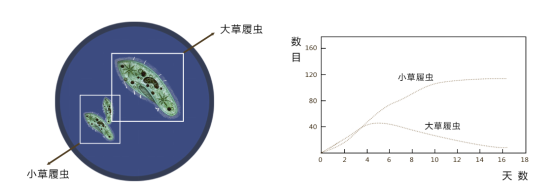
1934年,前苏联生态学家,Gause,用生态位相近的两个物种--原生动物双核小草履虫和大草履虫作为实验对象进行了实验。
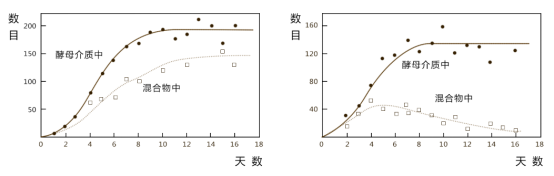
实验过程中,高斯发现:
(1)当分别在酵母介质中培养时,小草履虫比大草履虫增长快。
(2)当把两种加入同一培养器中时,小草履虫在混合物中占优势,最后大草履虫死亡消失。
用淡水硅藻和针秆藻一起培养后,也取得了同样的效果。
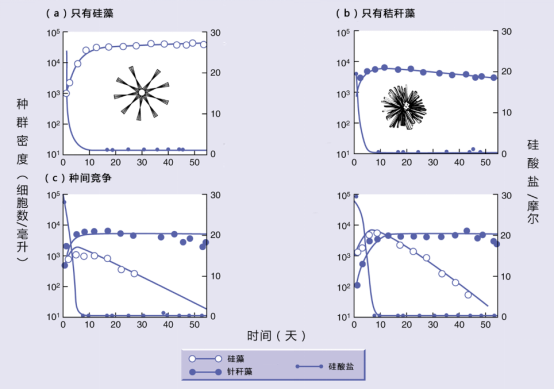
根据以上试验,我们看到了生态位相同的两个种不能在同一地区长期共存。因此,他提出假说
1.高斯假说
当两个物种利用同一种资源和空间时,产生的种间竞争现象表现为物种越相似,它们的生态位重叠就越多,竞争越激烈。这种种间竞争的情况称为高斯假说。
2.竞争排斥原理
近代人们又用竞争排斥原理来表示这种概念,即如果两个物种在稳定、均匀的环境中竞争,或者一物种胜利而另一物种被排除,或两物种达成共存。该假说表示竞争的结果。
共存只能在物种生态位分化的稳定,均匀环境中发生。如在以下情况下,则不会出现竞争排斥:
(1)不稳定的环境中,物种之间不能达到平衡;
(2)栖息在没有资源竞争的环境中的物种之间;
(3)在一物种被排挤掉之前,环境的改变使竞争方向改变。
![]()
(1)仅通过损耗有限的资源而个体不直接相互作用,即利用性竞争,;
(2)通过竞争个体间直接的相互作用,即干扰性竞争,
干扰性竞争最明显的例子是动物为了竞争领性或食物进行的打斗。干扰也可通过竞争者利用毒物来进行。在干扰性相互作用中“失败者”适合度的降低可能由于受伤,可能导致死亡,也可能由于缺乏可获资源。
![]()
种内竞争和种间竞争都时常很不均等的影响竞争者,一个个体的竞争代价远高于另一个体。
1. 争夺竞争
种内资源分布可能非常平均,所以大部分个体得到相似数量的食物。在高密度下,没有个体可得到足够活命的食物,从而使种群崩溃,这种状况叫争夺竞争。
2.格斗竞争
在一些种类,高密度导致资源的不平均分布,叫做格斗竞争。
3.竞争释放
在缺乏竞争者时,物种会扩张其实际生态位,这种现象称竞争释放。
在新几内亚的大岛上,发现3种地鸽,每种都分别处于不同的生境。可在在只有一种地鸽的小岛上,该种地鸽三种生境都可使用。
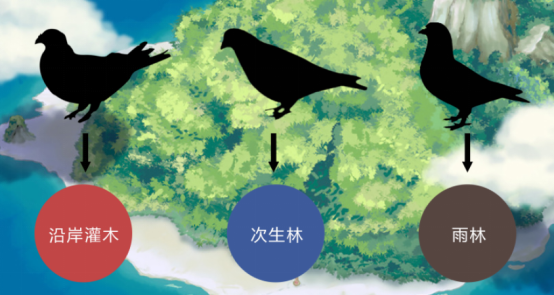
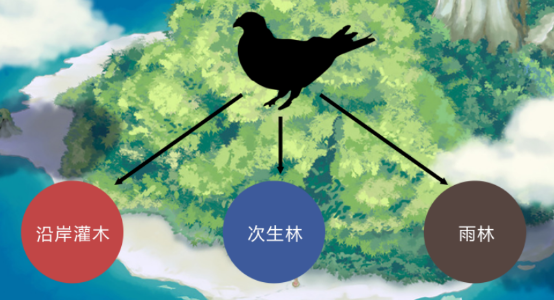
我们可以得出结论:竞争导致新几内亚大岛上地鸽的生态位分化,但在小岛上却发生了竞争释放。
4.性状替换
性状替换就是指两个亲缘关系密切的种类若在异域性分布中,它们的特征往往很相似,甚至难以区别。但在同域性分布中,它们之间的区别就明显,彼此之间必须出现明显的生态分离。
竞争导致的生态位收缩会导致形态形状发生变化,如:收获蚁的下颚大小与其他食种子的竞争蚂蚁的数量成负相关。这表明当其它蚂蚁参与食物竞争时,收获蚁只能转而食用一些体积更小的种子。
![]()
假设两个物种,当它们单独生长时增长曲线符合逻辑斯谛模型。
dN1 /dt = r1N1 (1-N1/K1)
dN2 /dt = r2N2 (1-N2/K2)
如果将这两个种放在一起时,它们之间会出现(资源、食物、空间的)竞争。
假设两个参数:α、 β
α表示在物种1的环境中,每存在一个物种2的个体,对于物种1产生的效应。
β表示在物种2的环境中,每存在一个物种1的个体,对于物种2产生的效应。
则有逻辑斯谛方程:
dN1 /dt = r1N1 (1-N1/K1-αN2/K1)
dN2 /dt = r2N2 (1-N2/K2-βN1/K2)
各参数的含义:
其中:
N1、N2:分别为两个物种的种群数量
K1、K2:分别为两个物种的环境容纳量
r1、r2 : 分别为两个物种的种群增长率
α、β:物种2对物种1的竞争系数,物种1对物种2的竞争系数。
![]()
从理论上看,物种1和物种2的竞争结果是由两个物种的竞争系数α、β和环境容量K1、K2共同决定的。
这里存在四中不同的情况:
1.α>K1/K2和β<K2/K1
1/K为种内竞争指标,
α/K1为种群2对1的种间竞争强度
β/K2为种群1对2的种间竞争强度
α>K1/K2,即:α/K1>1/K2
它表示:物种2中的每个个体对物种1的影响>物种2中每个个体对自身的影响
β<K2/K1,即:β/K2>1/K1
它表示:物种1中每个个体对物种2的影响<物种1中每个个体对自身的影响
这就使得物种1处于劣势,物种1被排斥,物种2 取胜
2.α<K1/K2和β>K2/K1
这个情况就跟第一种情况正好相反:
物种2中的每个个体对物种1的影响<物种2中每个个体对自身的影响。
物种1中每个个体对物种2的影响物种>1中每个个体对自身的影响
使得物种2处于劣势,物种2被排斥,物种1取胜
3.α>K1/K2或β>K2/K1
物种2中的每个个体对物种1的影响>物种2中每个个体对自身的影响
物种1中每个个体对物种2的影响>物种1中每个个体对自身的影响
此时:种间竞争>种内竞争。平衡点不稳定,两个物种都有可能获胜。
4.α<K1/K2和β<K2/K1
物种2中的每个个体对物种1的影响<物种2中每个个体对自身的影响
物种1中每个个体对物种2的影响<物种1中每个个体对自身的影响
此时:种间竞争<种内竞争。平衡点稳定,两个物种共存。
![]()
生态位是自然生态系统中一个种群在时间、空间上的位置及其与相关种群之间的功能关系。
通俗点说:生态位不仅要说明生物居住的场所,而且也要说明它要吃什么、被什么动物所吃、它们的活动时间、与其它生物的关系,以及它对生物群落发生的影响等所有方面。也就是说生态位是某一物种的个体与环境之间特定关系的总和。
1.生态位理论形成与发展
格里内尔(Grinell1917): 将生态位定义为物种栖息地再划分的空间单位,即:空间生态位。
埃尔顿(Elton1927):提出生态位是指物种在其群落中的地位和角色,强调与其他物种间的营养关系,即营养生态位。
哈钦森(Hutchinson1957): 提出生态位指在N维空间中的一个物种能够存活和增殖的范围,即:超体积和多维生态位。
奥德姆(Odum1971): 提出生态位不仅包括生物占有的物理空间,还包括了它在生物群落中的地位和角色,以及它们在各种环境变化梯度中的位置。
2.基础生态位与实际生态位
(1)基础生态位:
指生物群落中,某一物种所栖息的理论上最大空间,即没有种间竞争的种的生态位。
(2)实际生态位:
指当有竞争者时,必定使该物种只占据基础生态位的一部分,这一部分实际占有的生态空间称为实际生态位。 竞争种类越多,某物种占有的实际生态位越小。
实际生态位是生物之间的相互影响和物种自己对某些环境条件所具有选择性的共同结果
该概念对正确认识物种在自然选择进化过程中的作用,以及运用生态位理论指导人工群落建立中种群的配置等方面具有十分重要的意义。
3.生态位分类系统
(1)每一种生态因子对应着一种或一维特定的生态位,如光、温度、食物等生态位;
(2)按照生态元的类别,有基因、细胞、个体、物种、生态系统、城市、生物圈、地球生态位;
(3)根据竞争与否,分基础生态位和实际生态位,前者包括后者;
(4)生态位如果由生态元自身生产而形成的称为自产生态位,如鸟窝、人类建造的房、生态的粮食等;
(5)由其他生态元产生或自然存在、发生的生态位称非自产生态位
空间生态位:
表示划分环境空间单位和一个物种在环境中的地位。一个物种所占有的微环境。
营养生态位:
表示物种在生物群落中的地位与功能作用,强调的是物种与物种之间的营养关系。
这是一张植物根系资源分化,三种不同植物根在同一土壤区域同存。荨麻、印度锦葵、狗尾草
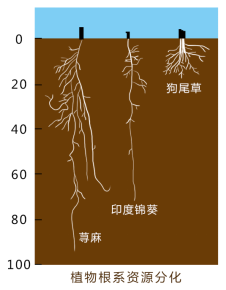
根据以上试验,我们看到了生态位相同的两个种不能在同一地区长期共存。若生活在同一地区,由于剧烈竞争,他们必然会出现栖息地、食物、活动时间或其他特征上的生态位分化。
![]()
1.生态位宽度
生态位宽度指一个有机体单位利用的各种各样不同资源的综合的幅度。是一种生物或生物类群所表现出来的资源利用的多样性。
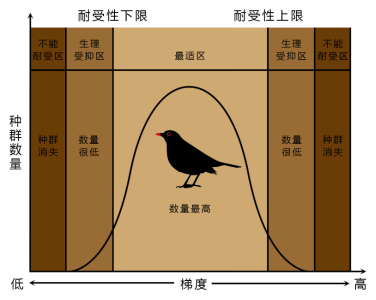
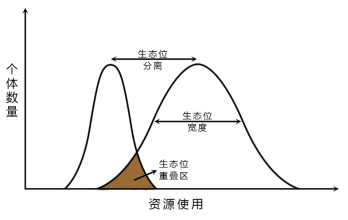
当可利用的资源少时,生态位宽度增加,促使生态位泛化。
当资源丰富时,可选择性大,生态位宽度减少,促使生态位特化。
2. 生态位重叠
生态位重叠指不同物种的生态位之间的重叠现象。或者说两个或更多的物种对资源位和资源状态共同利用。
生态位重叠是竞争的必要条件但并非绝对条件,而决定于资源状态。
当资源丰富,供应充足时,生态位重叠也不会发生种间竞争。
当资源贫乏,供应不足时,生态位稍有重叠即发生激烈的种间竞争。
3.生态位分离
生态位分离指两个物种在资源系列上利用资源的分离程度。又称竞争排斥原理或高斯。
它的原理是:如果许多物种占据一个特定的环境,他们要共同生活下去,必然要存在某种生态学差别,否则它们不能在相同的生态位内永久地共存。
4.生态位移动
生态位移动指种群对资源谱利用的变动。这是环境胁迫或者竞争的结果。
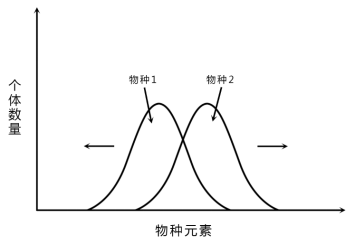
种内竞争使得两物种的生态位接近,种间竞争又促使两物种的生态位分离,或者一个种的灭亡
5.生态位分化与种内、种间关系
这是三个共存物种的资源利用曲线:
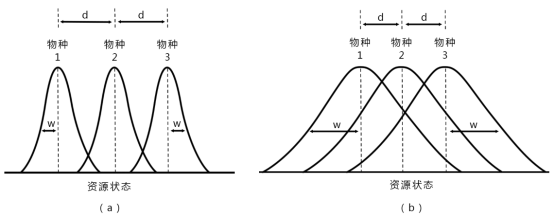
(a) 图生态位狭,相互重叠少;
(b) 图生态位宽,相互重叠多。
其中:d 为曲线峰值间的距离,w为曲线的标准差
当d>w时:种间竞争小,种内竞争强度大。
当w>d时:种内竞争小,种间竞争强度大。
极限相似性:指d/w = 1
![]()
(1) 一个稳定的群落中占据了相同生态位的两个物种,其中一个终究要灭亡;
(2)一个稳定的生物群落中,由于各种群在群落中具有各自的生态位,种群间能避免直接的竞争,从而保证了群落的稳定。
(3)群落是一个相互起作用,生态位分化的种群系统。这些种群在它们对群落的时间、空间和资源利用方面,以及相互作用的可能类型方面,都趋于互相补充而不是直接竞争。大家配合共同生活,更有效地利用环境资源,从而保证了群落在一个较长时间有较高的生长力,具有更大的稳定性。
(4)竞争可以导致多样性而不是灭绝,竞争在塑造生物群落的物种构成中发挥着主要作用。竞争排斥在自然开放系统中,很可能是例外而不是规律,因为,物种常常能够转换它们的功能生态位去避免竞争的有害效应。
![]()
生物多样性的分布格局和维持机制关键是物种的共存机制。
长期以来,生态位分化的思想在这一研究领域占据着主导地位。然而这一理论在解释热带雨林很高的物种多样性时遇到了困难。
由于生态位理论的不足,与实际情况似乎存在矛盾:
浮游生物悖论:环境条件非常均一的水体内共存着至少数十种浮游植物.
热带雨林:太高的物种多样性,无法确定每种植物的生态位。
由于生态位理论不能给这些群落学模式提供令人满意的解释。近年来中性理论应运而生。以美国生态学家Hubbell为代表的学者,认为:
(1)生态学上相同的物种可以实现共存,物种多度的变化是随机的,而非确定性的;
(2)共存的物种数量取决于物种分化和随机灭绝之间的平衡;
(3)群落内物种的相对多度随时间表现为随机振荡的波动。
生物的种间关系按性质可归并为两类。一是种间互助性的相互关系,如原始合作、共栖、共生等;二是种间对抗性的相互关系,如寄生、捕食、竞争等。![]()
1.捕食:
捕食是指一种生物以另一种生物个体(猎物)全部或部分为食的现象。
2.捕食的生态学意义:
(1)对猎物种群的数量和质量起着重要的调节作用。
(2)捕食者可以作为自然选择的力量对猎物的质量起一定的调节作用。
3、捕食者与猎物之间存在协同进化的关系:
(1)捕食者具有利用其猎物的适应性:比如锐齿、利爪、尖喙、毒牙等作为捕猎工具,诱饵追击、集体围猎。

(2)猎物具有逃避其捕食者的适应性:如保护色、警戒色、拟态、假死、集体抵御等。

![]()
草食作用是广义捕食的一种类型,其特点是被食者只有部分机体受损,留下的部分能够再生。
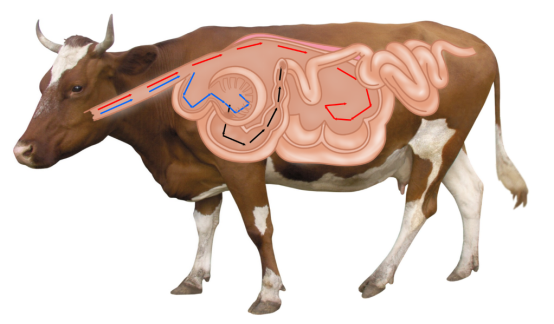
(1)食物的质量能影响捕食者消化和排泄系统的适应性:
由于植物内含有纤维素等物质使其难于消化,植食动物须将食物在消化管内停留较长时间才能彻底消化。这样就使植食动物胃内存留大量食物,降低了它的活动性。
(2)植物的补偿作用是指植物的枝叶在受损害后,自然落叶减少而整株的光合率可能增强。
(3)植物产生更多的刺,或化学分泌物都属于植物的防卫反应。
(4)同样,植物和草食动物也存在协同作用,在进化过程中,植物发展了防御机制,如有毒的次生物质,那么植食动物也产生相应的适应性,如形成特殊的酶进行解毒,或调整食草时间以避开植物的有毒化合物。
![]()
共生作用分为偏利共生和互利共生,偏利共生指共生中仅对一方有利,互利共生则是两物种相互有利的共居关系。
附生植物与被附生植物是一种典型的偏利共生关系,(如地衣、苔藓附在树皮上)

互利共生生物彼此间有直接的营养物质的交流,相互依赖、相互依存、双方有利。 如菌根、根瘤等则是互利共生关系

互利共生分为连体式互利共生非连体互利共生
菌根、根瘤这些属于连体式互利共生
犀牛和犀牛鸟、蝴蝶和花之间属于和非连体式互利共生。

