
Darwin在1859年的《物种起源》中详细地描述了繁殖与死亡现象相互作用,人为繁殖力是维持吴中延续的一个重要因子。
Lack于 1954年提出了动物的繁殖总是面对两种对立的进化选择:高生育力但无亲代抚育,低生育力但有亲代抚育。
R.H.MacArthur发展了以上两个理论,提出了r-k理论选择的自然选择理论。从而推进了生活史策略研究从定性描述走向定量分析的新阶段。
![]()
MacArthur和Wilson将生物按栖息环境和进化对策分为r-对策者和k-对策者,前者属于r-选择,后者属于k-选择。
自此,R选择和 K选择理论得到了正式认同 ,并且在以后的很多论文中被表述为竞争和扩散间的权衡。
1.r-选择
生活在条件严酷和不可预测环境中的种群,其死亡率通常与种群密度无关,种群内个体常把较多的能量用于生殖,而把较少的能量用于生长、代谢和增强自身竞争能力。
因此,我们把非密度制约性的自然选择常称之为r-选择。
例如,昆虫类:蚊子、苍蝇、蝗虫等昆虫,在秋冬季,天气寒冷,就几乎找不到它们,到了春夏季,天气转暖,他们就开始出来活动了。

2、k-选择
生活在条件优越和可预测环境中的种群,其死亡率通常由种群密度相关因素引起,生物间存在激烈竞争,种群内个体常把更多的能量用于生殖以外的其它各种活动。
因此,我们把密度制约性的自然选择常称之为k-选择。
例如:大型脊椎动物和高等级植物,就属于K对策。

![]()
r-对策者特征

快速发育,小型成体,后代数量多而个体小,繁殖能量分配高,世代周期短;
K-对策者特征

慢速发育,大型成体,后代数量少但体型大,繁殖能量分配低,世代周期长。
![]()
r-选择:
死亡率高,但r高能使种群迅速恢复,高扩散能力使其迅速离开不利环境,有利于建立新的种群和形成新的物种。
K-选择:
竞争能力强、数量稳定、大量死亡或导致生境退化的可能性小;由于r低,种群数量下降后恢复困难。
![]()
Grime认为:基本功能两对策模型的主要缺陷在于没有区分发生在不同类型的多年生植被中的两种完全不同的资源动态和优势度形成机制。
首先 ,在有持续资源补充的肥沃生境 ( 如冲积阶地 、 肥沃的草甸) 中,可通过迅速的资源捕获能力取得优势地位 。

第二,在贫瘠生境或生产力水平低的原生演替阶段,或演替后期成熟阶段,植被维持却并不依赖于优越的资源捕获能力,而是依赖于允许长寿组织残存并防御食草动物和病原体消耗已获得的资源的能力。

![]()
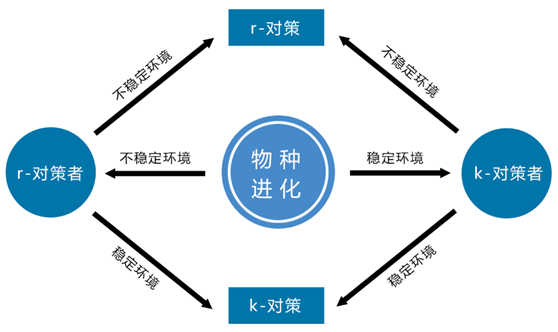
物种进化过程中,当遇到不稳定环境,不可预测的灾害比较多,只有r对策者能够应对而生存,当环境依然不稳定,向r对策进化;当环境稳定后,向k对策进化;
当环境稳定时,竞争较为激烈,只有k对策者能够应对,当环境越来越稳定,进化成k对策,当环境转向不稳定时,向r对策进化。
![]()
Grime提出:环境变化导致了植物生活史的发展,植物干扰强度和协迫强度,二个变量在选择中最重要,他以干扰强度和协迫强度组合为特征,建立了提出了CSR 理论。
理论内容:
CSR 理论中的,C代表竞争型植物; S代表耐胁迫植物; R代表杂草型植物
它的一个重要结论是 : 对于长期贫瘠或者具有频繁和严重干扰的生境 ,与其它有关适合度的要素相比 ,竞争的重要性降低。
C、 S 和 R 对策者是植物进化的极端情况 , 在中等生产力和中等干扰情况下 ,植物进化了次级对策。
Linhart(1974)年曾研究过水苦荬的两个种群,他们既是r-对策者又是K-对策者。
同样,竹子也是,当环境稳定时不开花结果;一但环境变的恶劣,缺少水和营养物质等,危协到生存时,就开花结种,采用种子的形式来保存遗传基因。

