
![]()
一、初级种群参数
初级种群参数包括出生率、死亡率、迁入和迁出等,我们先来看出生率。
1.出生率
出生率包括生理出生率和生态出生率
生理出生率:为在理想条件下所能达到的最大出生数量.
生态出生率:为种群在一定时期内特定条件下的实际出生数量,它是内外因素共同作用影响的结果.
影响出生率的因素有:
(1)性成熟速度
(2)每次产仔数
(3)每年生殖次数
(4)生殖年龄的长短.
2.死亡率
死亡率同样包括生理死亡率和生态死亡率。
生理死亡率:为最适条件下个体因衰老而死亡的数量,这里的种群死亡率降到最低。
生态死亡率:为一定条件下的实际死亡率。这里的许多个体死于各种生物或非生物影响的因素。
出生率和死亡率一般都以种群中每单位时间每1000个个体的出生或死亡数来表示。
3.迁入和迁出
迁入和迁出是种群变动的两个主要因子,它描述各地方种群之间进行基因交流的生态过程。
![]()
次级种群参数包括年龄结构和性比,对于一种生物个体,从出生到死亡都要经过出生、生长期、繁殖期、繁殖后期、死亡.
1.年龄结构及类型:
年龄结构:为不同年龄组的个体在种群内的比例和配置状况。种群的年龄结构与出生率死亡率密切相关。
当其他条件相等,种群中具有繁殖能力年龄的成体比例较大,种群的出生率就越高;而种群中缺乏繁殖能力的年老个体比例越大,种群的死亡率就越高。
种群有三种生态年龄,即3个年龄组:,分别为生殖前期、生殖期、生殖后期3种,
种群主要的年龄结构类型:分为增长型、稳定型和衰老型
增长型种群:基部宽,顶部狭。表示种群有大量幼体而老年个体较小,反映该比较年轻并且种群的出生率大于死亡率,是迅速增长的种群。
稳定型种群:大致呈钟型,从基部到顶部具有缓慢变化或大体相似的结构,说明幼年个体和中老年个体数量大致相等,出生率与死亡率大致相等,种群数量处于相对稳定状态。
下降型种群:呈壶型,基部比较狭、而顶部比较宽。表示种群中幼体比例很小而老体个体的比例较大,种群的死亡率大于出生率。说明种群数量趋于下降,为衰退种群。
从这张图中,我们可以看出肯尼亚、美国和澳大利亚三个国家的人口年龄结构。肯尼亚出生率很高,表现人口增长型;美国出生率适当,年龄结构合理,是稳定型;澳大利亚出生率偏低,年龄结构呈老龄化,是下降型。
种群的年龄分布体现种群存活、繁殖的历史,以及未来潜在的增长趋势,因此,研究种群的历史,便可预测种群的未来。
2.性比
性比是反映种群中雄性个体(♂)和雌性个体(♀)比例的参数。
受精卵的雄性个体和雌性个体比例,大致是1:1,这是第一性比,幼体成长到性成熟这段时间里,由于种种原因,雄性个体和雌性个体的比例变化,至个体开始性成熟为止,其的比例叫做第二性比,此后,还会有成熟的个体性比叫第三性比。
(1)性比对种群出生率的影响
性比对种群出生率有很大影响,例如:
一雄一雌繁殖的种群中,出生率由雌性个体所限制,我们假设,此种种群中种群总数为1000,其性比为6:4,那么,其能够进行繁殖的数量对就为400,而不是500。
一雄多雌繁殖的种群中,雌性个体比雄性个体多几倍,因此,出生率不受影响。
相反一雌多雄繁殖的种群中:雄性个体比雌性个体多几倍,因此,出生率必受到影响。
(2)性比变化的影响因素
环境条件:自然界中,大多数生物的自然种群内雌雄个体比率常为1:1;受环境因素影响,其比率可能会发生变化。如赤眼蜂在食物短缺时,雌性比例会下降.
发育时期:一定的发育时期内,生物的性比可能会发生变化,例如鳝鱼的前半生为雌性,后半生为雄性,其中间阶段则转变为雌雄间体。
(3)不同生育期的性比
一般,动物出生时雄性个体数量大于雌性个体数量。而到老的年龄组则会发生反转。
一、生命表对种群动态研究的作用
1. 生命表能综合评定各种群各年龄组的死亡率和寿命。
2. 生命表能计算出不同年龄组的个体比例情况;
3. 用生命表可以分析不同年份环境状况。
![]()
根据生命表所列数字的来源和类型,可将生命表分为动态生命表、静态生命表和综合生命表。下面我们以康内尔对藤壶的调查资料为例,说明不同生命表的编制方法。
1.生命表的内容
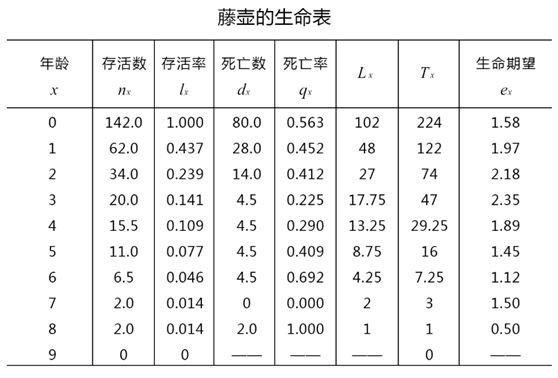
生命表有若干栏,每栏以符号代表,这些符号在生态学中已成为习惯用法,含义如下:
x=按年龄的分段age ;
nx=在x期开始时的存活数目;
lx=在x期开始时的存活分数;
lx = nx/ n0;
Lx是从x到x+1期的平均存活数。
Lx=(lx + lx+1 )/ 2x;
Tx: 进入x龄期的全部个体在进入x期以后的存活总个体一年值。
Tx =ΣLx ;
dx=从x到x+1的死亡数目
dx = nx – nx+1;
qx:从x到x+1的死亡率
qx= dx / nx;
ex=在x期开始时的平均生命期望或平均余年
ex = Tx / nx;
2. 动态生命表的编制
动态生命表是根据对同年出生的所有个体进行存活数目进行动态监察的资料而编制的。
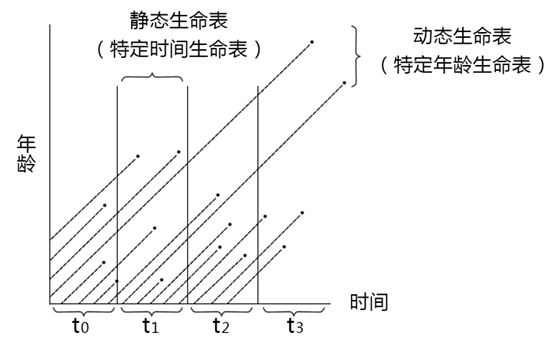
图中纵坐标表示年龄,横坐标表示时间。连续追踪t0,t21……时段中出生动物的存活就是动态生命表。因此,动态生命表也称为特定年龄生命表或水平生命表。
3. 静态生命表编制
静态生命表,是根据某一特定时间对种群作一年龄结构调查资料而编制的。
图中表示的根据t21时间所作年龄结构的生命表,就属于静态的,也称特定时间生命表或垂直生命表。
简单的生命表只能根据各年龄组的存活或死亡数据编制,综合生命表则能包括出生数据,从而估计种群的增长率。下面,我们就来看看,综合生命表的编制方法。
4. 综合生命表编制
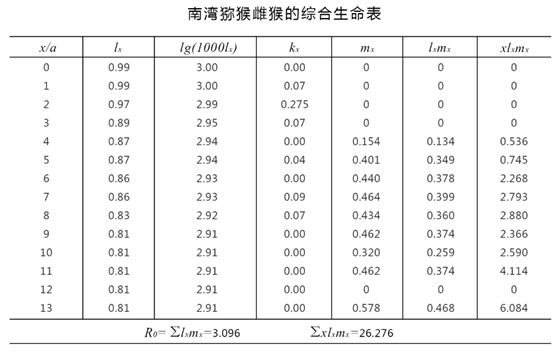
图中:
Lx为x年的存活数,kx为年龄组死亡率的指标,mx各年龄的出生率,RO净生殖率
由于综合生命表同时包括了存活率和出生率两方面数据,因此可以把各年龄的lx与mx相乘,并将其积累加起来,估计出一个非常有用的R0=∑lxmx值,称为净生殖率(net reproductive rate)。
因为R0是从各年龄组的lx和mx计算出的,它既包括该生命表所代表种群在该特定阶段中新出生的,也已扣除了死亡的,所以R0代表该种群(在生命表所包括特定时间中的)世代净增殖率。
利用生命表,我们就可以绘制出生物的存活曲线,从而能够对不同寿命的动物进行比较。
4.存活曲线
存活曲线是以存活数(nx)的对数对年龄(x)作图得到存活曲线。三种理想化的存活曲线模式用下图表示。
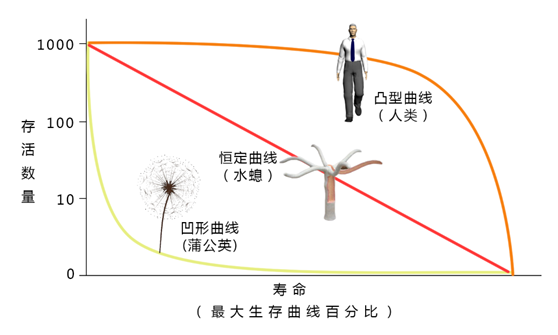
A型曲线:为凸型的存活曲线,表示种群在接近于生理寿命之前,只有个别的死亡,即几乎所有的个体都能达到生理寿命。死亡率直到末期才升高。如大型兽类和人类。
B型曲线:为呈对角线的存活曲线,表示个体各时期的死亡率是对等的。许多鸟类接近此型。
C型曲线:为凹型的存活曲线,表示幼体的死亡率很高,以后的死亡率低而稳定。如鱼类、两栖类、牡蛎等。
![]()
世代的净增殖率R0虽是有用的参数,但由于各种生物的平均世代时间并不相等,作种间比较时其可比性不强。种群增长率r值显得更有应用价值。
r值可按下式推导而得:
r=lnR0/T
其中T为世代时间,它是指种群中从母体出生到子代再产子的平均时间。用生命表资料可以估计出世代时间的近似值,即
T=(∑lxmxx)/(∑lxmx)
自然界的环境条件在不断变化着,当条件有利时,r值可能是正值,条件不利时可能变为负值。因此,长期观察某种群动态时,自然种群增长率r值的变化是很有用的指标。
为了进行比较;在实验室条件下,人们能排除不利的天气条件,提供理想的食物,排除捕食者和疾病,在这种人为的“不受限制”的条件下,就能观察到种群的最大的内禀增长率,rm。
因为实验条件并不一定是“最理想”的。所以由实验测定的rm值不会是固定不变的。正因为如此,人们能应用rm值为指标,测定某种生物种群的最适环境。

